Keywords
Wolbachia, Aedes aegypti, Yellow fever virus, vector competence
This article is included in the World Mosquito Program gateway.
Wolbachia, Aedes aegypti, Yellow fever virus, vector competence
Arboviruses impose a substantial disease burden on the human population1,2. Most recently, the Zika virus re-emerged in 2014, and unexpectedly caused serious congenital infections in pregnant women and Zika fetal syndrome in affected newborns in several American countries in 2016 and 20173. Chikungunya virus caused massive epidemics in the Americas in 2014 and still circulates in several countries4.
The yellow fever virus (YFV) is a member of the Flaviviridae family and transmitted by sylvan mosquitoes of the genus Haemagogus and Sabethes and Aedes aegypti in urban settings5–8. Monkeys are important reservoirs of YFV in sylvan environments. Encroachment by humans into environments where competent mosquito vectors and infected monkeys co-exist is the commonest reason for spillover of YFV transmission to human populations. Although the last reported cases of urban transmission in Brazil occurred in 1942, in 2016–2017, the country faced major outbreaks of the disease mainly in the states of Minas Gerais, Espírito Santo and Rio de Janeiro. In 2018, the epidemic also extended to São Paulo State9. According to the Brazilian Ministry of Health, from July 2017 to April 2018, there were 1,127 YFV cases with 328 deaths. Although the YFV vaccine is safe and effective, it does not always reach populations at greatest risk of infection and there is an acknowledged global shortage of vaccine supply10.
Recent studies have shown that anthropophilic mosquitoes, such as Aedes aegypti and Aedes albopictus, as well as Brazilian enzootic mosquitoes, such as Haemagogus leucocelaenus and Sabethes albiprivus, were highly susceptible to American and African YFV strains11,12. Therefore, the possible resurgence of urban epidemics of YFV in South America has to be constantly monitored by public health authorities13. Population control of Ae. aegypti mosquitoes using insecticides has been a mainstay of vector-borne disease control methods for decades but is undermined by widespread insecticide resistance. Wolbachia pipientis is a maternally transmitted bacterial endosymbiont and is naturally present in at least 40% of all insect species14. The World Mosquito Program is deploying Wolbachia pipientis as a self-sustaining disease control agent on the basis that Wolbachia reduces the transmission potential of Ae. aegypti mosquitoes for dengue15, Zika16 and chikungunya viruses17.
Here, we studied the ability of Wolbachia to suppress YFV infectivity in Ae. aegypti mosquitoes. Two virus isolates were used: one from a human clinical sample and another one of primate origin. We found that Wolbachia had a major impact on virus replication in mosquitoes and YFV transmission via saliva, as determined using a mouse model.
The first sample named M377_IV|Human|MinasGerais_PadreParaíso|2017-02-04 (YFV377H) was isolated from human serum, positive for YFV by RT-qPCR (CT = 28.95) in February, 2017 from Padre Paraíso city (northeast of Minas Gerais state). The other sample named M127_IV|Primate|MinasGerais_NovaLima|2018-01-15 (YFV127P) was isolated from the liver of a non-human primate found dead in January 2018, in Nova Lima city, in the center-south of Minas Gerais state, positive for YF via RT-qPCR (CT = 17.19). Sequencing of both isolates was performed and is described below. Viral isolation was confirmed by two methodologies: indirect immunofluorescence (IFA) and real-time PCR. IFA was performed with a monoclonal YFV antibody donated by Evandro Chagas Institute (Arbovirology and Hemorrhagic Fevers Section) and conjugated goat anti-mouse IgG labeled with fluorescein FITC (MP Biomedicals). Images were obtained using an Olympus microscope model BX51 with DP72 camera and DP-2BSW software. Viral molecular confirmation was performed using RNA extracted from the culture supernatant of each isolate, followed by amplification of the genetic material as described below in the viral detection section. For mosquito infections, the YFV isolates were replicated in C636 cells (Ae. albopictus) cultured in Leibovitz 15 medium (Gibco) supplemented with 10% fetal bovine serum (FBS) (Gibco) for 5 days at 28°C. Viral load was confirmed by RT-qPCR and later through plaque assays (PFU) in VERO cells (CCL81) grown in DMEM medium (Gibco) and 3% Carboxymethylcellulose (Sigma) supplemented with 2% FBS (Gibco) at 37°C and 5% CO218.
Viral RNA was isolated from 200µL of each sample using MagNA Pure 96 (Roche) following manufacturer’s recommendations. To confirm the viral presence in isolates, RT-qPCR was performed, according to Domingo et al. 201219.
A real-time nanopore sequencing strategy with previously developed primers20, was applied to both RT-qPCR-positive samples. For these samples, extracted RNA was converted to cDNA using GoScript™ Reverse Transcriptase (Promega) and random hexamer priming. Whole-genome amplification by multiplex PCR was attempted using GoTaq® qPCR Master Mix (Promega), the 500bp sequencing primer scheme and 35 cycles using the adapted protocol20. Electrophoresis (2% agarose gel) was used to confirm the expected bands and to purify the specific amplicons using Invitrogen™ E-Gel™ SizeSelect, followed by quantification using fluorimetry with the Qubit dsDNA High Sensitivity assay on the Qubit 3.0 instrument (Life Technologies).
Template was amplified with end point PCR to increase template concentration following the Ion Plus Fragment Library Kit recommendation and PCR products were cleaned-up using AmpureXP purification beads (Beckman Coulter). Emulsion PCR was performed to amplify the library using Ion PGM™ Hi-Q™ View OT2 Kit (Thermo Fisher Scientific) and the Ion OneTouch 2 system (Thermo Fisher Scientific). Ion Sphere particles (ISPs) were enriched using the Ion OneTouch ES (Thermo Fisher Scientific). Enriched ISPs were sequenced using the Ion Torrent Personal Genome Machine (PGM) and the Ion PGM Hi-Q Sequencing kit (Thermo Fisher Scientific), with the Ion 314 chip. All procedures above followed manufacturer’s instructions.
Consensus genome sequences from fastq file were produced by alignment of two-direction reads by using a reference YFV genome. Quality control on raw sequence data have been performed using FastQC21. Bowtie 2 was used for mapping reads to a reference using Galaxy22. Only positions with ≥ 20× genome coverage were used to produce consensus sequences. Regions with lower coverage and those in primer-binding regions were masked with N characters.
In order to identify the origin of the YFV genome from the samples, we performed a maximum likelihood (ML) phylogenetic analysis using the newly two nucleotide sequences recovered in this study plus 125 reference YFV complete genome sequences from each different genotype (South American I n=84; South American II n=2; West African n=23; East African n=16) already published in peer-reviewed journals, for which sampling year and geographic location is available. Full details of the reference sequences used are provided in Extended data: Table S1.
Consensus sequences were aligned using MAFFT v.723. Maximum likelihood phylogenetic trees were estimated using IqTree24 under a GTR + Γ4 nucleotide substitution model. Statistical support for phylogenetic nodes was estimated using a bootstrap approach (100 replicates).
The phylogenetic signal has been investigated with the likelihood mapping method by analyzing groups of four sequences, randomly chosen, called quartets. Likelihood mapping analyses was performed with the program TREE-PUZZLE by analyzing 10,000 random quartets25.
Wild type Aedes aegypti mosquitoes collected in the neighborhood of Urca, Rio de Janeiro-RJ, Brazil in 2018 were reared in the laboratory for five generations and confirmed for the absence of Wolbachia (WT). Wolbachia wMel strain-containing mosquitoes (wMel +) were obtained from the colony maintained by the World Mosquito Program (WMP) Brazil laboratories in Belo Horizonte, which is backcrossed every five generations with Urca male mosquitoes. They were reared in a controlled environment at 27 ± 2°C and 60 ± 10% relative humidity. Four to six days-old female mosquitoes were starved for 20 to 24 hours and subsequently offered YFV virus culture supernatant mixed with washed human red blood cells (RBCs) (2:1 ratio). The viral titer offered to mosquitoes was 4 × 105 PFU/mL for YFV377H and 1.4 × 106 PFU/mL for YFV127P. RBCs were washed three times for removal of potential YFV vaccine antibodies. Mosquitoes were allowed to feed for one hour and then, engorged females were selected and maintained in triple containment, under BSL-2 conditions. Sucrose solution (10%) was offered ad libitum during the extrinsic incubation period. Viral load was analyzed at 7, 14 and 21 days post feeding (dpf), via RT-qPCR. Additionally, a subset of mosquitoes (at 7dpf) received an extra blood meal and were collected at 14dpf, when Wolbachia density and viral load was determined. The blood used in the infective feedings was obtained from a blood bank (Hemominas) through an agreement signed between both institutions (OF.GPO/CCO-Nr224/16). As a laboratory routine each blood bag is previously tested for dengue, Zika, chikungunya, mayaro and yellow fever, through RT-qPCR to rule out any cross-infection that could interfere with the results.
In order to check the ability of mosquitoes to transmit the virus, saliva samples from infected mosquitoes were individually collected at 14 dpf. After removal of legs and wings, mosquitoes had their proboscis introduced into 10 μL tips, containing 50% Fetal Bovine Serum (FBS) (Gibco) and 30% sugar solution and allowed to salivate for 30 minutes. Mosquitoes and solution containing the saliva were stored at -70°C until RNA extraction of the heads/thoraces and/or nanoinjection of the saliva into naive mosquitoes (WT). Saliva samples were injected into WT mosquitoes, after 2 to 4 days of emergence. Each mosquito received 276 nL and were kept for 5 days before whole body RNA extraction, followed by RT-qPCR.
In vivo experiments were conducted using type I interferon receptor deficient mice (A129−/−), SV129 background. A129-/- originally from The Jackson Laboratories (reference 010830) were obtained from Biotério de Matrizes da Universidade de São Paulo (USP) and kept under specific pathogen-free conditions at Immunopharmacology Lab at UFMG. Mice were housed in filtered-cages of 28x13x16 cm with autoclaved food and water available ad libitum on ventilated shelves (Alesco). A maximum of 4 mice were kept per cage. Mice were housed under standard conditions with controlled temperature (18–23 degrees) humidity (40–60%) and 12/12h dark light cycle. Sample sizes for in vivo studies were determined using the G*Power 3.1 software package. In each experiment we used 4 mice on YFV377H or YFV127P groups and 6 mice per group on saliva YFV 377H or 127P infected mosquitoes (WT or wMel+) groups. Mice from the same litter were added to either mock- or YFV infected groups, or test or control groups as appropriate. No randomization protocol was utilized. For most of the experiments, no blinding was involved except for body weight and hind paw swelling analysis. Bioanalysis from viral loads and cell count assay experiments was blinded. Groups were divided by codenames on the day of euthanasia. Different researchers performed the euthanasia or analyzed the data. Each experiment was replicated twice and all attempts at replication were successful. For the experiments, adult A129-/- mice (7 to 9 weeks old, 20-22g) were inoculated with 1 × 104 PFU with either YFV377H or YFV127P viruses’ strains or with a pool of saliva samples (n=2) either from the WT or wMel+ groups via subcutaneous (intraplantar) route/50μl paw (right hind paw). Morbidity parameters such as body weight loss, total and differential counts of blood leukocytes and paw edema were evaluated daily. Total cell counts were carried out in Trypan blue-stained cells in a Neubauer chamber and differential cell counts on blood smears stained with May-Grunwald-Giemsa using standard morphological criteria. Paw edema was assessed by measuring paw swelling using a pachymeter. Finally, viable viral loads and viral RNA were analyzed in plasma and different tissues of mice upon saliva inoculation, as shown below.
All animal experiments involving YFV infection and Wolbachia saliva inoculation were conducted following the ethical and animal welfare regulations of the Brazilian Government (law 11794/2008). The experimental protocol was approved by the Committee on Animal Ethics of the Universidade Federal de Minas Gerais (CEUA/UFMG, permit protocol no. 84/2018). All surgeries were performed under ketamine/xylazine anesthesia and all efforts were made to minimize animal suffering. Studies with YFV were conducted under biosafety level 2 (BSL-2) containment at Immunopharmacology Lab from Instituto de Ciências Biológicas (ICB) at Federal University of Minas Gerais.
Detection of viral particles on infected mosquitoes and mice samples were performed through quantitative real-time PCR (RT-qPCR) using LightCycler® Multiplex RNA Virus Master (Roche), according to the previously published protocol26. RNA extractions were performed following manufacturer's protocols. Mosquito samples were processed through the High Pure Viral Nucleic Acid kit (Roche), mice tissue samples (liver, spleen) were extracted with Trizol (Invitrogen), whereas mice lymph node samples were isolated with the QIAamp® Viral RNA kit (Qiagen). Multiplex reactions were performed with primers and probes described in Table 1. Reactions were performed on a Lightcycler96 real-time PCR machine (Roche) with the following program: first step at 50°C for 10 min for reverse transcription, 95°C for 30 sec for inactivation and initial denaturation and 95°C for 5 sec followed by 60°C for 30 sec for 40 cycles. The reaction volume was 10 μL (5× RT-PCR Reaction Mix (Roche), 200× RT-enzyme solution (Roche), 2.5 μM each primer (IDT) and 2 μM YF (target yellow fever) probe (IDT) and 1 μM WSPTM2 (target wMel-specific) probe and 0.7 μM RPS 17S (target Ae. aegypti ribosomal S17) probe. For mouse samples, only the YFV probe was used. A fraction (1/20) of the total isolated RNA was used in the reactions. Viable viral loads were quantified by titration assay in permissive Vero cells as described in Costa et al., 201227.
| Sequence 5’→3’ | Reference | |
|---|---|---|
| YFV Forward | GCTAATTGAGGTGYATTGGTCTGC | 19 |
| YFV Reverse | CTGCTAATCGCTCAAMGAACG | |
| YFV Probe | FAM/ATCGAGTTG/ZEN/CTAGGCAATAAACAC/3lABkFQ | |
| WSPTM2 Forward | CATTGGTGTTGGTGTTGGTG | 15 |
| WSPTM2 Reverse | ACACCAGCTTTTACTTGACCAG | |
| WSPTM2 Probe | CY5/TCCTTTGGA/TAO/ACCCGCTGTGAATGA/3lAbRQSp | |
| RPS17 S Forward | TCCGTGGTATCTCCATCAAGCT | 29 |
| RPS 17S Reverse | CACTTCCGGCACGTAGTTGTC | |
| RPS17 S Probe | HEX/CAGGAGGAG/ZEN/GAACGTGAGCGCAG/3lABkFQ |
All statistical analyses were performed on Prism (Graphpad Version 7.04). Initially the D'Agostino and Person normality test was performed. Wolbachia density data as well as viral load were compared using the non-parametric Mann-Whitney test. Statistical analyzes for the mouse data were performed with ANOVA one-way test. The significance level was set for p values less than 0.05.
Two plasma samples (one human and one from a non-human primate) were isolated from the diagnostic service of Fundação Ezequiel Dias, the State Reference Laboratory of Minas Gerais, Brazil. Viral isolation was confirmed by indirect immunofluorescence (IFA), showing the typical signal of fluorescence for both isolates (Figure 1B and C). Both samples were successfully sequenced with PGM (Personal Genome Machine) technology with adapted overlapping multiplex PCR protocol, as shown in Table 2. The phylogenetic analysis showed that the isolates obtained from the two samples (M377_IV and M127_IV) belonged to the South American genotype I and clustered closely with strong bootstrap support (>90%) with the recent sequences, isolated in Minas Gerais, from the current outbreak (Figure 2)28.
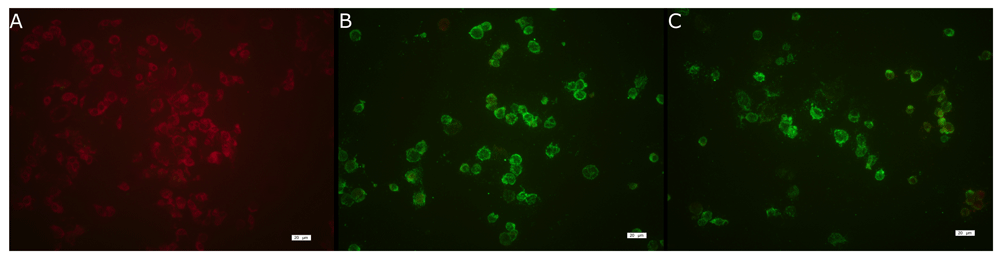
(A) Control cells without virus, (B) cells infected with YFV 377 H and (C) cells with YFV127 P. Green fluorescence depicts YFV in cells marked with a monoclonal YFV antibody conjugated goat anti-mouse IgG labeled with fluorescein FITC.
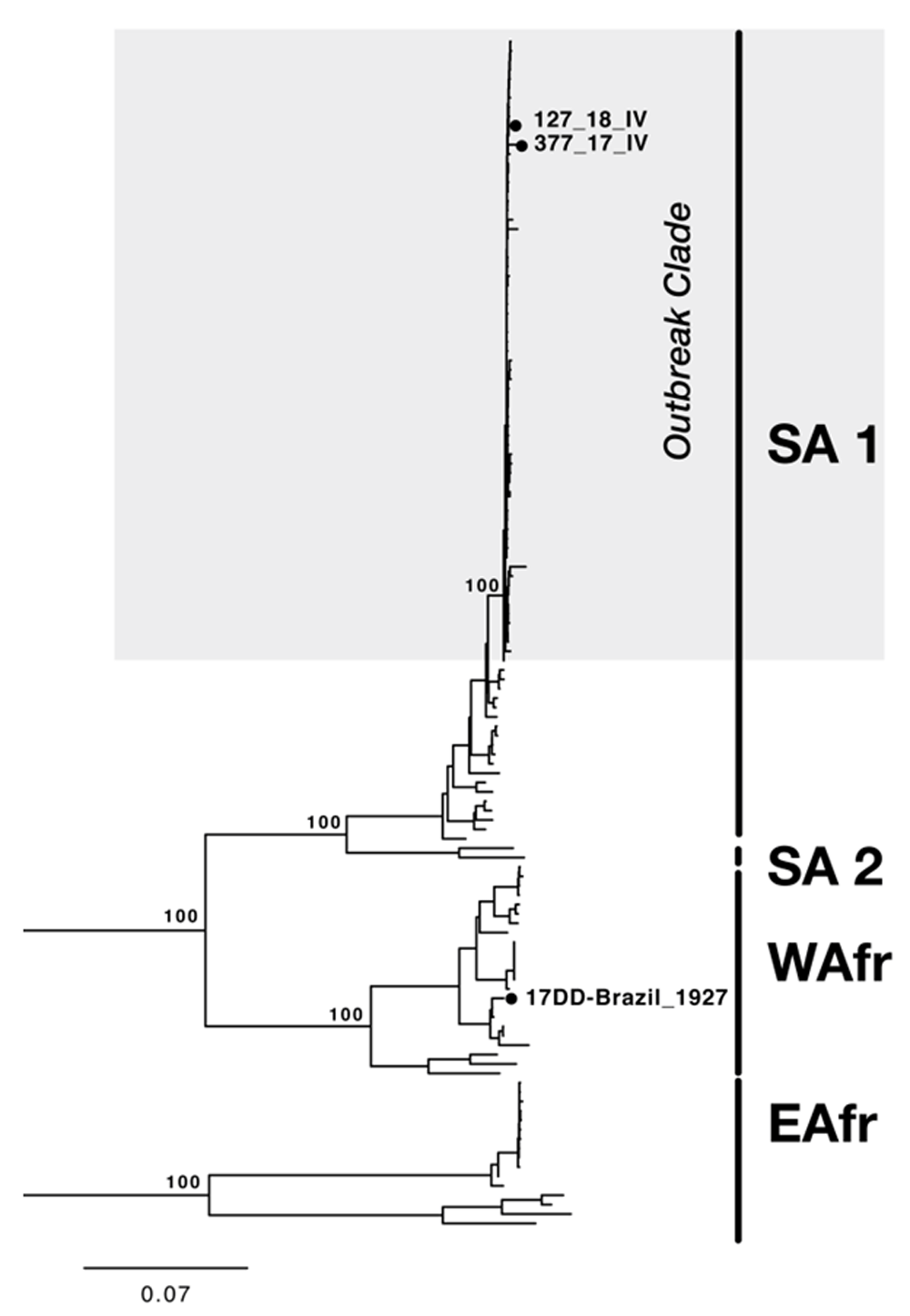
ML showing the two newly genomes belongs to South American I (SA1) genotype. SA2, WAfr, and EAfr indicate the South America II, West Africa, and East Africa genotypes, respectively. The scale bar is in units of substitutions per site (s/s). Node labels indicate bootstrap support values.17DD, the vaccine strain used in Brazil.
Absolute quantification of Wolbachia in mosquitoes were analyzed in the heads + thoraces of Wolbachia-positive mosquitoes (wMel +) after challenge with YFV. There was no difference in Wolbachia density among heads and thoraces, collected at 7 or 14 days post feeding (dpf), as shown in Figure 3A. However, Wolbachia density presented a slight reduction at 21dpf, which was statistically significant in relation to 14dpf (p = 0.0062, Mann Whitney). The median at 14dpf was 2.04 × 106 copies per head/thorax whereas at 21dpf, it decreased to 1.37 × 106.
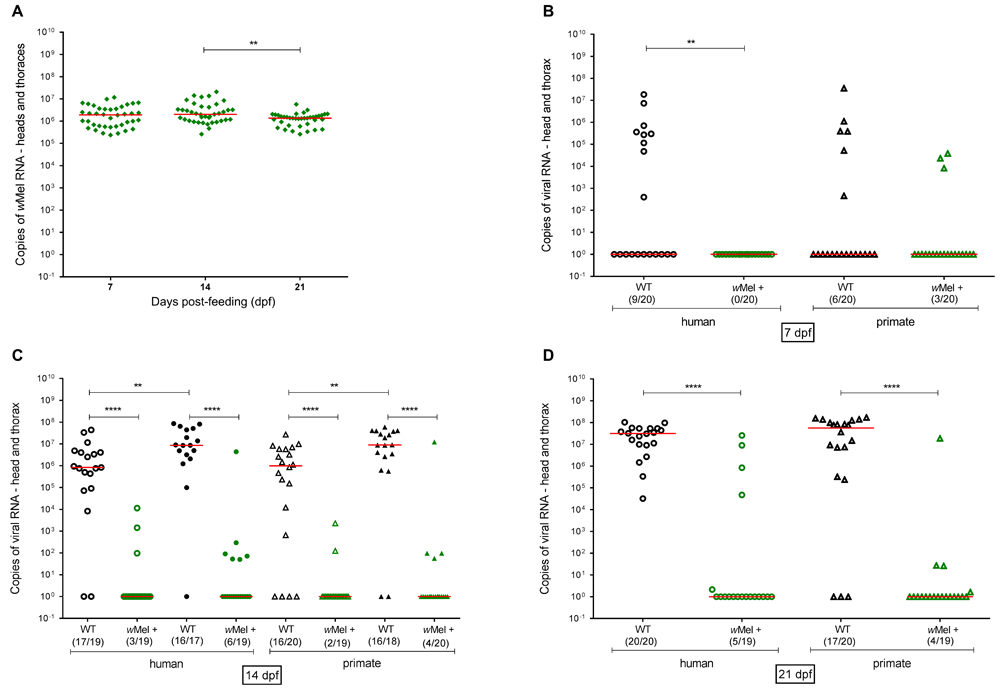
Wild type (WT) or positive (wMel +) were orally infected with two YFV isolates and virus dissemination in mosquitoes was analyzed at different times post infection. (A) YFV infected mosquitoes’ heads and thoraces were analyzed for Wolbachia density at different times post-infection through real time RTq-PCR, based on a Wolbachia standard curve. (B) Analysis of 7dpf p=0.0012, (C) 14dpf **** p<0.0001 and YFV Human ** p=0.0050, YFV Primate ** p=0.0046 and (D) 21 dpf **** p=0.0001. Empty black circles and triangles are WT mosquitoes, whereas empty green circles and triangles depict mosquitoes with wMel +. Black filled circles and triangles are mosquitoes that received a second blood meal. The red line indicates the median YFV copies. Red lines indicate the median wMel copies. ** p=0.0062; analysis performed through the Mann-Whitney U test.
In mosquitoes without Wolbachia (WT) the prevalence of YFV infection of heads + thoraces was 30–45% at 7dpf, and 80-89% at 14dpf. For those mosquitoes that received a 2nd blood meal, the prevalence was 89 to 94% at 14dpf and 85 to 100% at 21dpf. There was no significant difference between infection rates resulting from the human or primate virus isolates (Figure 3). In heads + thoraces of Wolbachia-positive mosquitoes (wMel +) the infection rate ranged from 0 to 15% at 7dpf, 11 to 16% at 14dpf, 20 to 32% at 14dpf when mosquitoes received a second blood meal, and 20 to 25% at 21dpf (Figure 3). Again, there was no major difference between viral isolates.
The infection rate observed at 7dpf was low for both viral isolates (Figure 3B). At day 7, the presence of Wolbachia was already associated with a marked decrease in viral titers in mosquitoes (Figure 3B). At 14dpf, there was a significant increase in the number of viral copies in WT mosquitoes (Figure 3C). Further increase on viral load was observed when mosquitoes received a second blood meal 7 days after the infective meal and were analyzed at 14 dpf. This increase was statistically significant for both isolates (p <0.01, Mann Whitney). This may have been due to the fact that the second blood supplied extra important nutrients for viral replication. At 21dpf, the infection reached 100% for the human isolate with a median of 3.15 × 107 viral copies. For the primate isolate, although the infection rate was lower (85%), the viral load was higher with a median of 5.61 × 107 viral copies per head/thoraces. Regardless of the strain of virus used, viral loads were remarkable lower in presence of Wolbachia at all time points (Figure 3B–D). In addition, there was no increase in viral load in wMel + mosquitoes after supplying a second blood meal (Figure 3C).
Next, we evaluated the ability of orally infected mosquitoes to transmit the virus. We first collected saliva from infected mosquitoes at 14 dpf, from both groups of mosquitoes and virus isolates. We then injected a number of saliva samples into eight naïve (WT) mosquitoes and, after five days, we checked whether those mosquitoes became infected through RT-qPCR, demonstrating that a particular saliva was infectious. As shown in Figure 4, when saliva samples originated from wMel + mosquitoes, no mosquitoes became infected. This assay shows, indirectly, the potential of Wolbachia to completely abrogate YFV transmission potential of Ae. aegypti mosquitoes. Nevertheless, saliva originating from WT mosquitoes was able to infect 20% of the naïve-injected mosquitoes.
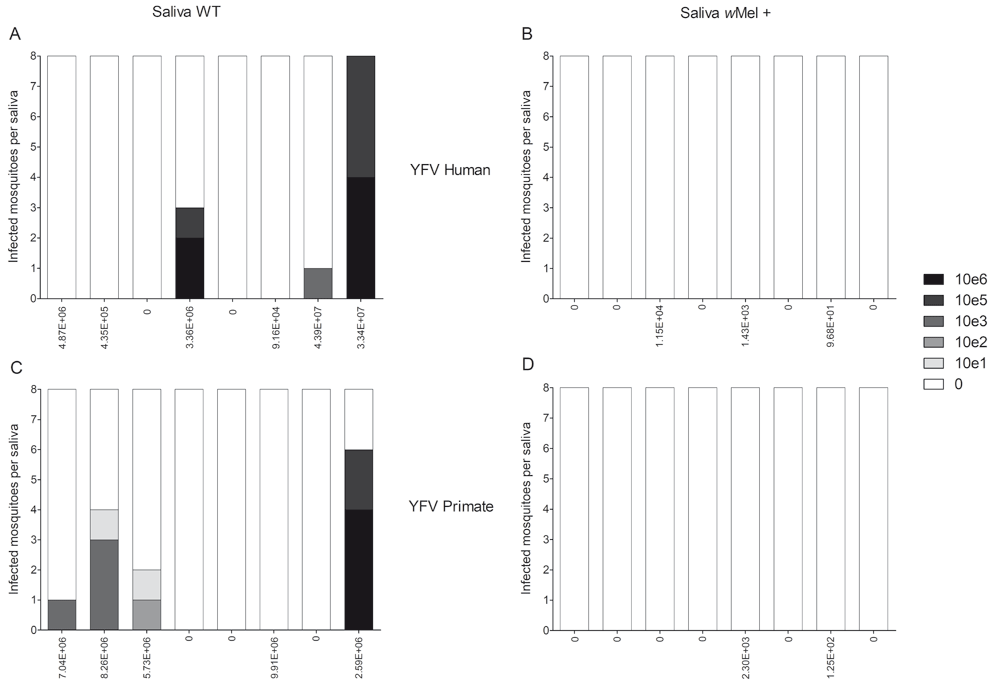
Saliva from both groups of infected mosquitoes were collected at 14 dpf. Individual saliva samples (WT or wMel +) were injected into eight naïve (WT) mosquitoes (bars) and, after five days, these injected mosquitoes were analyzed. (A) Mosquitoes injected with wMel+ mosquito saliva or (B) WT mosquitoes, challenged with human virus. (C) Mosquitoes injected with wMel+ mosquito saliva or (D) WT mosquitoes, challenged with primate virus. Values below each bar depicts the viral load of each mosquito head and thorax which donated that saliva. Positive mosquitoes were quantified through qPCR and the grey-scale represents the number of YFV copies (0 to 106 copies), per mosquito.
Similar experiments were performed by injecting saliva samples from either the WT or wMel + groups into 4-week-old A129-/- mice, which are susceptible to arboviral infections30,31. Results showed that there was no major impact on clinical and laboratory parameters, which is consistent with the relatively low number of viable virus injected (Figure 5A–D). However, there were viable viruses, as assessed by plaque assay, recovered from the paw of mice inoculated with saliva from WT mosquitoes. Indeed, there was culturable virus when both P (primate) and H (human) strains were used. In contrast, none of the samples from the wMel + groups were positive on the plaque assay (Figure 5E–H). Consistently with the mosquito saliva findings above, there were higher number of viral RNA copies in draining lymphnode and liver from mice injected with WT saliva than mice inoculated with wMel + saliva (Figure 5 I–K). Virus isolated from the primate (YFV127P) showed greater presence in liver while the human strain (YFV377H) was more localized at the lymphoid tissue (Figure 5).
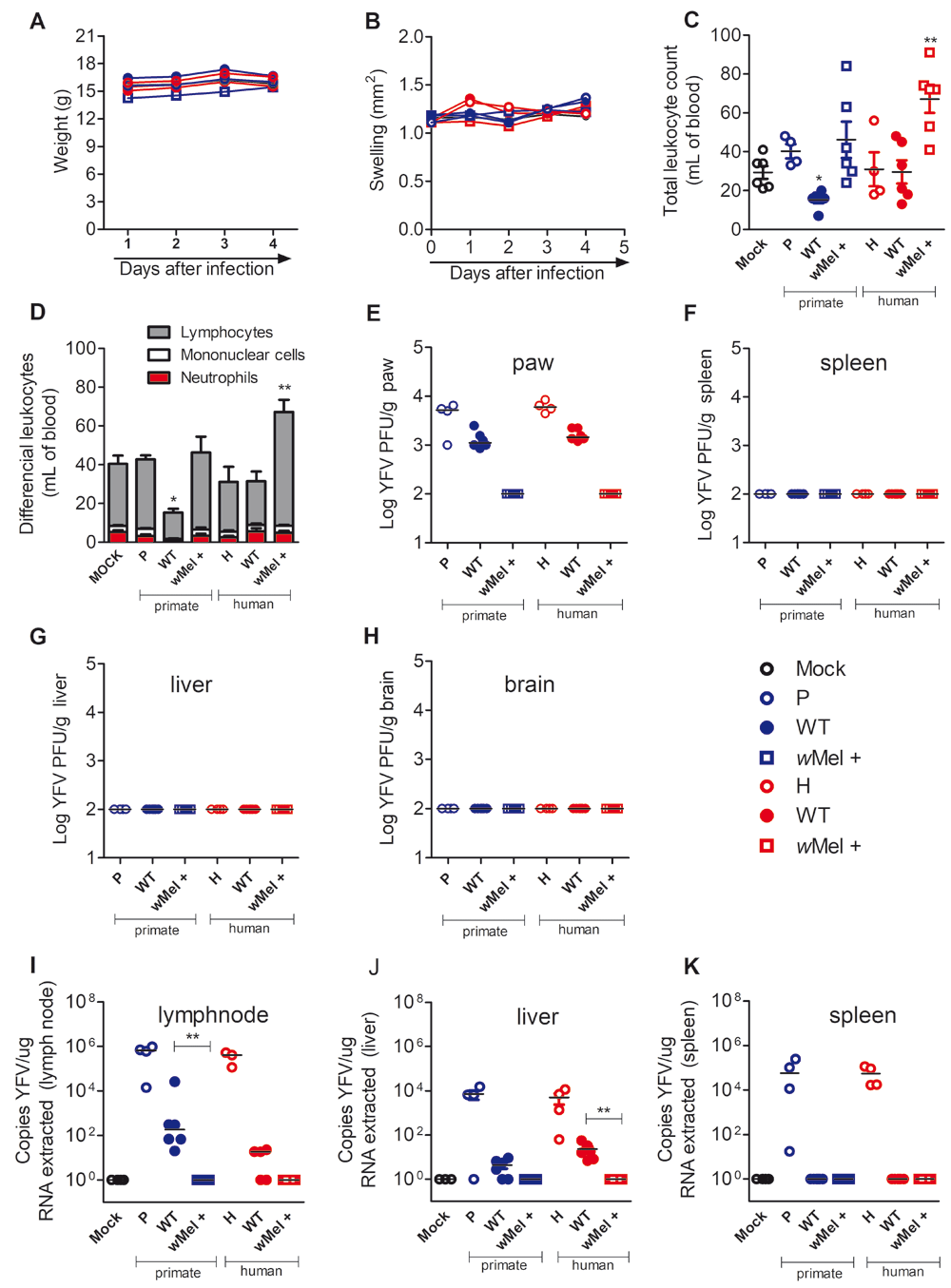
A129-/- mice were inoculated with 1 × 104 PFU of YFV primate (empty blue circles) and human YFV (empty red circles) or with a pool of saliva from wild tipe (WT) YFV primate (full blue circles), WT YFV human (full red circles), Wolbachia-positive (wMel +) YFV primate (empty blue squares) and Wolbachia-positive YFV human (empty red squares) previously infected with YFV via intraplantar route (50 μl/paw). Control mice (MOCK group) received 50 μl of PBS solution (empty black circle). (A) Body weight analysis shown as body weight (g) of mice. (B) Paw volume measured daily and shown as swelling (mm2). On day 4 post-infection mice were euthanized and the following analysis performed. (C–D) Total and differential leukocyte counts in the blood. (E–H) Viable viral loads recovered from paw (E), spleen (F), liver (G) and brain (H) by plaque assay in Vero cells. Results are shown as Log PFU/g of tissue. (I–K) Viral RNA copies recovered from popliteal lymph node (I), liver (J) and spleen (K) by RT-qPCR. Data was presented as meanSEM or median (n=4 mice for MOCK, n=6 mice for WT P, wMel + P, WT H and wMel + H groups and n=4 for YFV P and YFV H, one-way anova).
Collectively these results suggest that Wolbachia-positive mosquitoes can efficiently suppress YFV replication and reduce virus transmission through saliva.
The ability of Wolbachia to reduce the susceptibility of Ae. aegypti to disseminated arbovirus infection has been repeatedly demonstrated for dengue15, Zika16, chikungunya17, West Nile32 and mayaro virus26. We have shown that wMel was able to significantly reduce the infectivity of YFV to mosquitoes, independently of the source of the virus (both human and primate). Previously, it has been shown that two strains of Wolbachia (wMelPop and wMel) were able to significantly reduce YFV mosquito infection, although with virus isolated from human cases from Nigeria and Bolivia, in 1987 and 1999, respectively33. Here we evaluated the effect of Wolbachia (wMel strain) towards two recently isolated yellow fever viruses, originating from the 2017–2018 outbreaks in Brazil. The yellow fever virus isolates used here have different origins, one originating from a non-human primate found in the city of Nova Lima and another originated from a human case in the city of Padre Paraíso, both in the state of Minas Gerais. Although these cities are located more than 500 km apart, they belong to the same genotype. Besides working with recently isolated virus from human and primate sources, the difference in the present study refers to the way mosquitoes have been infected. Furthermore, this study was performed with orally infected mosquitoes, which is closer to natural conditions, in comparison to the previous study which infected mosquitoes through thorax injection, in order to improve mosquito infection34.
The use of Wolbachia as an arbovirus control strategy has been developed by the not-for-profit initiative, the World Mosquito Program. The approach offers the prospect of a natural and sustainable method for arbovirus control34–37. The impact towards reduction of arbovirus has been analyzed38,39 and early indication of positive effect has been recently reported40. In Brazil, WMP is expanding its coverage into Rio de Janeiro and Niterói municipalities and epidemiological studies in order to determine arbovirus reduction is underway.
The blocking ability conferred by Wolbachia has been directly related to the density of the bacterium within main mosquito tissues such as midgut and/or salivary glands15,41, where viruses replicate to further produce infectious particles42. In our study, and as observed by Pereira et al., 201826, the density of Wolbachia was constant at 7 or 14 days after virus exposure. However, there was a reduction of wMel + density at 21dpf, which did not impact the blocking ability towards the virus (Figure 3). The variation on the density (or titer) of Wolbachia within the host has been previously observed, which could be related to the aging of the host41.
In the present study, the presence of Wolbachia in mosquitoes greatly reduced YFV infection, except for 7dpf, when the infection rate was low in all groups. Further effect of Wolbachia towards YFV was verified when individually collected mosquito saliva was injected into naïve mosquitoes or into a susceptible mice strain and their infectivity was analyzed. This first technique has been widely used by our group and others16,26,43, and it is a robust proxy of the potential of individual saliva towards virus transmission. When the source of saliva came from Wolbachia-positive mosquitoes, there was no infection in any injected mosquito. Through projection of these results into natural conditions, the YFV transmission could be greatly reduced, as previously modeled for dengue virus38.
Another interesting fact of this work was the increase in viral load observed after the second blood feeding in WT mosquitoes. This same fact was not observed in wMel + mosquitoes. This shows that the blocking ability of Wolbachia persists even after the addition of extra blood nutrients (through a second blood meal) and that its blocking effect occurs within 7 days after infection. Interestingly, in our experiments, the overall infectivity in mosquitoes was not high, even in control (no Wolbachia) mosquitoes. This shows the reduced vector competence of natural Brazilian Ae. aegypti populations, which could explain why most of the cases reported on the recent outbreaks in Brazil were in proximity to green areas of parks and forests, where natural YFV mosquito vectors such as Haemagogus and Sabethes are easily found11,12,44.
Our results show that the presence of wMel strain of Wolbachia in mosquitoes has the potential to greatly reduce the transmission potential of Ae. aegypti for YFV. It is important for public health agencies of arbovirus endemic countries to have constant awareness of the potential of Ae. aegypti to become an urban vector for yellow fever once again6,45. If that becomes reality, Wolbachia-infected mosquitoes could be a powerful tool for YFV control, along with the currently applied vaccination program10,46. Integration of complementary strategies are the best solution for arbovirus control.
The data underlying Figure 3, Figure 4 and Figure 5, as well as viral sequencing data is available from Open Science Framework, https://doi.org/10.17605/OSF.IO/PUZ6947.
Data are available under the terms of the Creative Commons Zero “No rights reserved” data waiver (CC0 1.0 Public domain dedication).
Genome sequences generated in this study are publicly available in GenBank database: M377_IV|Human|MinasGerais_PadreParaiso|2017-02-04: accession number, MK249065; M127_IV|Primate|MinasGerais_NovaLima|2018-01-15: accession number, MK249066.
| Views | Downloads | |
|---|---|---|
| Gates Open Research | - | - |
|
PubMed Central
Data from PMC are received and updated monthly.
|
- | - |
Provide sufficient details of any financial or non-financial competing interests to enable users to assess whether your comments might lead a reasonable person to question your impartiality. Consider the following examples, but note that this is not an exhaustive list:
Sign up for content alerts and receive a weekly or monthly email with all newly published articles
Register with Gates Open Research
Already registered? Sign in
If you are a previous or current Gates grant holder, sign up for information about developments, publishing and publications from Gates Open Research.
We'll keep you updated on any major new updates to Gates Open Research
The email address should be the one you originally registered with F1000.
You registered with F1000 via Google, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Google account password, please click here.
You registered with F1000 via Facebook, so we cannot reset your password.
To sign in, please click here.
If you still need help with your Facebook account password, please click here.
If your email address is registered with us, we will email you instructions to reset your password.
If you think you should have received this email but it has not arrived, please check your spam filters and/or contact for further assistance.
{kind=link}
{kind=link}
Comments on this article Comments (0)